
Absorción de CO2 por los cultivos más representativos de la Región de Murcia
24 de noviembre de 2010
Las plantas tienen la capacidad de captar el CO2 atmosférico y mediante procesos fotosintéticos metabolizarlo para la obtención de azúcares y otros compuestos que requieren para el normal desarrollo de su ciclo vital (Fig 1. Fotosíntesis, (1)). En general, se puede concluir que, las plantas, a través de la fotosíntesis, extraen el carbono de la atmósfera (en forma de CO2) y lo convierten en biomasa. La biomasa al descomponerse se convierte en parte del suelo (en forma de humus) o en CO2 (a través de la respiración de los microorganismos que procesan la biomasa (Fig. 1 (2)).

En la actualidad, el exceso de CO2 modifica en balance final del ciclo de carbono descrito anteriormente, influyendo de manera decisiva sobre las condiciones climáticas. Por una parte se produciría una captación del CO2 de la atmósfera por parte de las plantas a través de la fotosíntesis. Por otra parte, la respiración de las plantas, las quemas y las talas para usos agrícolas incrementan en la atmósfera la concentración de emisiones de CO2, lo que unido a una tasa de deforestación alta y a las escasas medidas de reforestación aplicadas altera el balance entre emisión y captación. De esa manera la concentración de CO2 en la atmósfera va aumentando. Estas emisiones netas del sector agrícola y forestal se suman a las emisiones de CO2 que se generan al quemar combustibles fósiles en los sectores de transporte y generación de energía (Fig 1 (4)).
Sumideros de carbono
La captación de CO2 por los ecosistemas vegetales terrestres constituye un componente importante en el balance global de Carbono (C). A escala mundial se considera que la biosfera terrestre fija cerca de 2.000.000 toneladas/año (Unesa, 2005). Este valor es el resultante de la pequeña diferencia entre la absorción fotosintética de CO2 y las pérdidas por respiración, por descomposición de la materia orgánica y por perturbaciones de diferente naturaleza. A este valor le se denomina producción neta de la biosfera (PNB), y es la cantidad que a largo plazo queda almacenada en el sumidero.
El CO2 secuestrado por las plantas es el resultado de las diferencias entre el CO2 atmosférico absorbido durante el proceso de la fotosíntesis y el CO2 emitido por la atmosfera durante la respiración. Esta diferencia es convertida en biomasa y suele oscilar entre el 45-50 % del peso seco de la planta. Por lo tanto, mientras el crecimiento sea alto, la vegetación natural y los cultivos agrícolas se convierten en los sumideros de carbono. Teniendo esto en cuenta, la agricultura se puede convertir en un mecanismo efectivo para mitigar el incremento del CO2 atmosférico.
La fotosíntesis

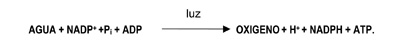
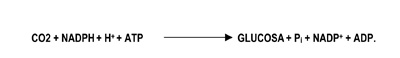
En las reacciones de oscuridad, el CO2 de la atmósfera (o del agua en organismos fotosintéticos acuáticos/marinos) es capturado y reducido por la adición de hidrógeno (H+) para la formación de carbohidratos [(CH2O)]. La incorporación del dióxido de carbono en compuestos orgánicos, se conoce como fijación o asimilación del carbono. La energía usada en el proceso proviene de la primera fase de la fotosíntesis. Los seres vivos no pueden utilizar directamente la energía luminosa, sin embargo, a través de una serie de reacciones fotoquímicas, la pueden almacenar en la energía de los enlaces C-C de carbohidratos, que, más tarde, será liberada mediante los procesos respiratorios u otros procesos metabólicos.
Plantas C-3: Se caracterizan por mantener los estomas abiertos durante el día para permitir la fijación de CO2, lo que provoca una pérdida de agua por transpiración, de forma continua. Ante el riesgo de deshidratación ocasionado por un estrés ambiental, estas plantas producen un cierre estomático que provoca una gran disminución de la fotosíntesis.
Plantas C-4: Se caracterizan por tener los estomas abiertos de día. Como poseen intermediarios de bombeo de CO2 en la célula, pueden permitirse un cierre de estomas imprevisto, siendo factible la continuidad del proceso fotosintético, gracias al reservorio de CO2.
Plantas CAM: Estomas abiertos por la noche. Las pérdidas de agua por transpiración se reducen enormemente. También poseen reservorio de CO2, con lo cual también pueden cerrar estomas sin que ello conlleve una disminución fotosintética.
Las propiedades de las plantas C-4 y CAM les permiten una supervivencia en entornos con déficit hídrico.

Efecto de los estreses ambientales sobre la fijación de CO2
Dado el carácter fuertemente desecante de la atmósfera, el control de las pérdidas de agua ha sido siempre un aspecto clave para las plantas. Por una parte, el flujo de agua a través de una planta debe ser suficiente para mantener la nutrición y la incorporación de CO2. Y por otra, como la asimilación y la transpiración están estrechamente ligadas en casi todas las plantas, la disponibilidad de agua impone un límite máximo a la productividad (desarrollo) (Steudle and Peterson, 1998). Al mismo tiempo, para evitar la desecación de las partes aéreas, el flujo de agua que entra en la planta por las raíces ha de compensar la salida de agua por las hojas. Dado que los procesos fisiológicos son extremadamente sensibles al déficit hídrico, la conservación del agua para mantener potenciales hídricos razonablemente altos suele ser el principal problema en las zonas con climas cálidos y escasez de precipitaciones.
Se estima que la fijación de CO2 se verá incrementada en los próximos 60 años debido al aumento en la temperatura. Se espera que la fijación de CO2 se incremente el 1% por cada °C en regiones donde la temperatura media anual es de 30 °C y el 10% en regiones donde la temperatura media anual es de 10 °C. Las tasas fotosintéticas subirían un 25-75%, en las plantas de fotosíntesis C3 (las más comunes en latitudes medias y altas), al duplicarse la concentración de CO2. Los datos son menos concluyentes en el caso de las plantas cuya modalidad fotosintética es la C4, típica de lugares cálidos, siendo los intervalos de respuesta desde 0% hasta un 10-25% de incremento (Unesa, 2005).
La agricultura de la Región de Murcia tiene un peso medio en el PIB. Se trata de una de las agriculturas más rentables de España y de Europa debido a su alta productividad, que está muy por encima de la media nacional. La agricultura murciana está orientada a la exportación, lo que implica el desarrollo de una industria agroalimentaria potente y una necesidad de red de comunicaciones de primer orden. Si a todo esto añadimos las actividades indirectas que genera la agricultura, ésta adquiere una importancia notable dentro de la economía regional.
Las excelentes condiciones climáticas de nuestra región, junto al uso intensivo de los avances de la revolución verde aumentan notablemente la rentabilidad de nuestra agricultura. La escasez de agua en la región se ha convertido en un factor limitante provocando que, en la actualidad, los regadíos dependan de las aguas subterráneas (que contienen un gran contenido salino debido a la sobreexplotación así como a la intrusión del agua del mar), ya que el trasvase Tajo-Segura se ha quedado insuficiente para cubrir las necesidades de la región.
Los productos hortícolas, en sus múltiples variedades, son los productos más importantes en la agricultura murciana: tomate, lechuga, pimiento, alcachofa, etc., aunque también tienen gran relevancia los cítricos (en especial el limón) y los cereales, seguido del viñedo, junto a otros productos arborícolas con una alta rentabilidad, como el almendro, el melocotonero, el ciruelo, etc.
En general, la explotación forestal tiene poca importancia económica, y ocupa poca superficie de la región. Se localizada, mayoritariamente, en las comarcas montañosas y no cubren las necesidades de la región. El bosque autóctono ha sufrido importantes ataques antrópicos, por lo que las principales especies forestales son los pinos de repoblación y los chopos en las riberas de los ríos.
Unas buenas prácticas agrícolas o una gestión sostenible de las fincas (no dejar el suelo descubierto, utilizar cantidades exactas de abono en el momento y en el lugar exacto, no quemar cosechas y reducir el arado) supondrían dejar de emitir millones de toneladas de gases de efecto invernadero. Por esto, se establece un código de buenas prácticas agrarias relativas a la protección del suelo, al mantenimiento de la materia orgánica y de la estructura del suelo y a la conservación de los hábitats y del paisaje, incluida la protección de los pastos permanentes. Este cambio en el modelo agronómico podría suponer un balance positivo de CO2 en las superficies agrícolas. Con un conocimiento y gestión adecuados, este sector puede contribuir a la mitigación de estos gases mediante la adecuación del laboreo de la tierra, la promoción de la producción ecológica y el uso más eficiente de recursos en la maquinaria agrícola, convirtiéndose finalmente en eficiente
Por lo tanto, como parte de este proyecto, en este trabajo de investigación, se ha determinado la tasa de captación de CO2 anual por los cultivos más representativos de la Región de Murcia basados en los datos de producción de biomasa y su contenido de carbono. Se han elegido aquellos cuyas hectáreas totales de regadío supongan una superficie mayor de 1000 Ha. Se ha calculado la captación de carbono por plantas individuales, teniendo en cuenta únicamente la biomasa anual. De esta forma, se ha estimado el carbono fijado total o el CO2.
Material y métodos
Posteriormente, se introdujeron en una estufa de aire caliente a 70 °C hasta peso constante para determinar el peso seco. El proceso de secado puede variar dependiendo de la humedad y peso total de la muestra. Una vez obtenido el peso seco de cada muestra de las diferentes partes de la planta, se molieron en un molinillo de laboratorio, IKA modelo A10. El resultado fue muy homogéneo con partículas de 5 a 7mm de diámetro. El total de carbono se analizó tal y como se describe posteriormente.


Para el trasporte del material de muestreo desde la finca de muestreo hasta la cooperativa, se utilizaron palops de 30kg y una transpaleta motorizada En la cooperativa se pesaron las muestras por separado en una báscula de suelo compuesta por un puente de pesado de chapa de acero acanalada antideslizante, cuatro células de pesado móviles y un terminal de manejo.
El muestreo de los frutos se realizó de los frutos recolectados en la campaña. Una muestra representativa de los frutos se trasladó al laboratorio para la obtención del peso seco y el contenido de carbono total, tal y como se describe en los apartados anteriores. El total de cosecha de fruto se estimó de la media obtenida en todos los árboles en la parcela de muestreo.

Una muestra representativa de cada órgano, junto a muestras de fruto, se recolectaron para su determinación de peso seco en el laboratorio como se ha descrito anteriormente.
Para la expresión del Carbono total capturado por árbol y por año, se consideró que la biomasa de hoja se renueva cada 3 años y que el peso total de la parte aérea y de la raíz está en una relación del 70/30 respectivamente de la biomasa total del árbol (Morgan et al., 2006). Las medidas se realizaron como se describe a continuación

El total de carbono se analizó en sub-muestras (alrededor de 2-3 mg PS) de hojas, tallos, frutos y raíces con un analizador de NC-Thermo Finnigan 1112 EA analizador elemental (Thermo Finnigan-, Milán, Italia).

Resultados: Estimación de carbono y captación de CO2 en plantas herbáceas
En las plantas de tomate (Tabla 1) se observa un mayor contenido de carbono y captación de CO2 que en las de pimiento (Tabla 2) ya que el tomate presenta una mayor biomasa que el pimiento. Sin embargo, cuando se calcula el total de carbono por hectárea, las diferencias entre estos dos cultivos se reducen debido a que la densidad del cultivo de pimiento es mayor (2,2 plantas m-2) frente a tomate (2 plantas m-2). En la actualidad coexisten una alta cantidad de distintas variedades de tomate en la región, y distintos tipos de cultivo. Para el estudio se eligió el tomate de ensalada (variedad Corvey) y cultivo en suelo
 en tomate")
 en pimiento")
 en sandía")
 en melón")
 en las distintas variedades de lechuga...")
 en dos variedades de bróculi...")
 en plantas de coliflor...")
 en plantas de alcachofa...")
Estimación de CO2 y contenido de carbono en cereales



Estimación de carbono y captación de CO2 en árboles frutales
Las tablas recogen los valores medios de biomasa y captación de CO2 totales a partir del porcentaje de carbono de cada una de las fracciones de la planta en las que han sido divididas. El total de carbono se ha determinado en la planta completa, teniendo en cuenta la producción anual de fruto y el crecimiento anual de la planta.
En la tabla 12 se muestran los datos correspondientes al análisis del albaricoquero donde se observa un mayor contenido de carbono y captación de CO2 por árbol que en el resto de los frutales de hueso, pero teniendo en cuenta que la densidad de plantación del albaricoquero es la mitad que la del resto de frutales de hueso, sería el melocotonero el que mayores índices por superficie obtendría (tabla 14). De hecho, si tuviéramos en cuenta solamente el contenido en carbono y la captación de CO2 por unidad de superficie, el albaricoquero sería la especie con menores índices seguido por el ciruelo (tabla 13). Los valores más altos se producen para el melocotonero y nectarino (tabla14 y 15). Hay que tener en cuenta que el ciruelo es el frutal de hueso con menor peso seco (biomasa) de los cuatro analizados, lo que indicaría una mayor capacidad de captación de CO2 y de acumulación de carbono.
Los datos obtenidos para parras de uva de mesa (tabla16) muestran que a pesar de tener aproximadamente la mitad de peso seco que el nectarino, obtiene valores similares en los índices de acumulación de carbono por unidad de superficie. Por otra parte, cuando se comparan los índices de acumulación de C y de captación de CO2 por parra con los datos obtenidos de los frutales de hueso muestra valores muy inferiores (hasta un 75% de disminución si comparamos con el albaricoquero).





Estimación de CO2 en cítricos
En el caso de los cítricos es el limonero (tabla 17) el que consigue valores superiores, no sólo al compararlos con los del resto de los cítricos sino también cuando se compara con el resto de especies arbóreas. En este caso, el limonero muestra mejores índices de captación y acumulación tanto por árbol (debido a que tiene mayor biomasa que el resto de especies) como por superficie. En general parece el cultivo con mayor capacidad de fijación de CO2. El naranjo (tabla18) presenta valores muy inferiores al limonero pero similares en general a los frutos de hueso, mientras que el mandarino mostró valores inferiores al resto (tabla19).



")

Discusión
En este trabajo se han presentado los datos de captación de CO2 por planta para poder comparar entre especies agrícolas, pero para una mayor consideración de los balances en el cálculo total hay que tener en cuenta los resultados por hectárea junto al conocimiento de la densidad de plantación.
En general, de los datos obtenidos de este trabajo podemos afirmar que de los cultivos hortícolas analizados, la alcachofa es el más eficiente en su captación de CO2 seguido de tomate y sandía (Figura 1). Sin embargo, cuando se analizan los resultados por unidad de superficie, es la coliflor el cultivo más eficiente y la alcachofa pasa a ser de los menos eficientes junto a la sandía y melón. Los cereales cuando se analizan por planta individual son muy eficientes en la fijación de CO2 superando a todos los valores obtenidos en las plantas hortícolas. Sin embargo, cuando se analizan por superficie, los resultados descienden a valores muy bajos.
Entre todas las especies arbóreas analizadas en este trabajo es el limonero el que mostró mayor índice de captación de CO2, tanto en función de la superficie como por árbol. En este caso el factor más relevante para la captación del CO2 es el gran desarrollo vegetativo que alcanza el limonero a lo largo de su vida, convirtiéndose en árboles más frondosos, con mayor superficie foliar y, por lo tanto, con mayor capacidad de captación de CO2. En la agricultura moderna, el naranjo y el mandarino se cultivan con un porte mucho menor que el limonero. Sin embargo, a pesar de que su marco de plantación es menor, sus índices de captación de CO2 resultan muy inferiores a los de éste, e, incluso, a los del melocotón y el nectarino, teniendo éstos un peso seco inferior al naranjo. En este caso, el factor limitante para la captación de CO2 por la planta es el marco de plantación utilizado para el correcto cultivo del naranjo.
Los cálculos del presente trabajo, indican también las posibles direcciones políticas si se quiere incrementar el índice de captura del CO2 atmosférico. Por un lado, se ha de propiciar una mayor extensión de cubierta vegetal con agricultura en zonas donde la cubierta natural es escasa y por otro, hay que tener en cuenta que mayores aportes hídricos van a determinar un incremento en la biomasa agrícola. En este sentido, el clima semi-desertico de gran parte de la Región de Murcia produce altos incrementos en la evapotranspiración y por consiguiente mayores requerimientos hídricos (Cubasch et al., 2001).
Como se ha podido desprender del trabajo presentado, dependemos de las plantas para contrarrestar el efecto invernadero. Por lo tanto, la solución al cambio climático pasa necesariamente por la conservación de la mayor cantidad posible de zonas con vegetación. Debemos optimizar su capacidad de captación con la mejora de las prácticas agronómicas y con la utilización de los subproductos. Por otro lado, la alta capacidad de adaptación que poseen las plantas y que les ha permitido resistir grandes cambios a lo largo de miles de millones de años, ha de utilizarse como base para estudios científicos que nos permitan evaluar la situación de nuestra agricultura en las condiciones climáticas futuras.
Por todo ello, los resultados encontrados inciden sobre la necesidad de conservar los sistemas agrícolas de nuestra Región con mayores aportes hídricos que determinarán un incremento en la biomasa agrícola y por lo tanto una mayor captación de CO2 atmosférico. Todo ello con el compromiso de reutilizar los subproductos para obtener energía, fertilizantes e incluso el agua retenida en los órganos o tejidos de la planta no utilizados.
Los autores agradecen a las siguientes empresas o agrupaciones, su disposición a la toma de muestras, ayuda técnica y asesoramiento en todo momento de este estudio: Langmead Farms, Finca experimental del Cebas-CSIC, José Peñalver Fernández, CDTA El Mirador, Morte Quiles, Frutas Esther, Patricio Peñalver Aznar, Frutas Torero, Aproexpo y Fecoam.
- Båth, B. (2000) Matching the availability of N mineralisation from green-manure crops with the N-demand of field vegetables, PhD Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden.
- Biodisol.com (2009) http://www.biodisol.com/biocombustibles/investigadores-la-upv-crean-un-proceso-para-reutilizar-los-desechos-de-los-citricos-usados-para-zumos-energias-renovables-biocombustibles-investigacion-e-innovacion/
- Blanco-Roldán, G., Cuevas, S. (2002). La importancia de la maquinaria para la poda y manejo de restos. Vida Rural. 1 de Febrero de 2002, 56-58.
- Bouwman, A. F. (1994) Direct Emission of Nitrous Oxide from Agricultural Soils (Report No. 773004004, National Institute of Public Health and Environmental rotection, Bilthoven, the Netherlands)
- Brady, N.C. and Weil, R.R. (2004). Elements of the Nature and Properties of Soils, 2/E. Ed. Pearson Prentice Hall, N.J.
- Cubasch U, Meehl GA, Boer GJ et al. (2001) Projections of future climate change. In: Climate Change 2001: The Scientific Basis.Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change (eds Houghton JT), pp. 525582. Cambridge University Press, Cambridge.
- Dumas Y., Dadomo M., Di Lucca G., Grolier P. (2003) Effects of environmental factors and agricultural techniques on antioxidant content of tomatoes, J. Sci. Food Agric. 83, 369382.
- Gunnarsson S. (2003) Optimisation of N release Influence of plant material chemical composition on C and N mineralization, PhD Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden.
- IPCC (Intergovernmental Panel on Climate Change). Good Practice Guidance for Land Use, Land-Use Change and Forestry. (2003). Edited by Jim Penman, Michael Gytarsky, Taka Hiraishi, Thelma Krug, Dina Kruger, Riitta Pipatti, Leandro Buendia, Kyoko Miwa, Todd Ngara, Kiyoto Tanabe and Fabian Wagner.
- Kimball B.A., Kobayashi K., Bindi M. (2002) Responses of agricultural crops to free-air CO2 enrichment, Adv. Agron. 77, 293368.
- Kirchmann H., Johnston A. E. J., Bergström L. F. (2002) Possibilities for reducing nitrate leaching from agricultural land, Ambio 31, 404408
- Lal R. (1997). Residue management, conservation tillage and soil restoration for mitigating greenhouse effect by CO2-enrichment. Soil Tillage Res 43:81107
- Mäder P., Fliessbach A., Dubois D., Gunst L., Fried P., Niggli U. (2002) Soil fertility and biodiversity in organic farming, Science 296, 16941697.
- Martínez-Ballesta, M.C., Lopez-Perez, L. Muries, B, Muñoz-Azcarate, O., Carvajal, M. (2009) Climate change and plant water balance. The role of aquaporins. Sustainable Agricultural Reviews (E. Lichtfouse, Ed.) Vol 2, 71-89.
- Matson, P.A., Naylor, R. Ortiz-Monasterio I. (1999) Integration of environmental, agronomic, and economic aspects of fertilizer management. Science 280: 112-115.
- Morgan, K.T., Scholberg J.M.S., Obreza T.A. Wheaton T.A. Size, (2006) Biomass, and Nitrogen Relationships with Sweet Orange Tree Growth J. Amer. Soc. Hort. Sci. 131(1):149156..
- Robertson GP, Paul EA and Harwood R, (2000).Greenhouse gases in intensive agriculture: contributions of individual gases to the radiative forcing of the atmosphere. Science 289:19221925
- Sofo, A., Nuzzo, V., Palese, A.M., Xiloyannis, C., Celano, G, . Zukowsky P., Dichio. B. (2005) Net CO2 storage in Mediterranean olive and peach orchards. Sci. Hortic. 107:1724.
- Solomon, S., D. Qin, M. Manning, R.B. Alley, T. Berntsen, N.L. Bindoff, Z. Chen, A. Chidthaisong, J.M. Gregory, G.C. Hegerl, M. Heimann, B. Hewitson, B.J. Hoskins, F. Joos, J. Jouzel, V. Kattsov, U. Lohmann, T. Matsuno, M. Molina, N. Nicholls, J. Overpeck, G. Raga, V. Ramaswamy, J. Ren, M. Rusticucci, R. Somerville, T.F. Stocker, P. Whetton, R.A. Wood y D. Wratt. (2007): Technical Summary. In: Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge y New York: Solomon, S., D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor y H.L. Miller ed. Pp, 25.
- Steudle E., Peterson C. (1998) How does water get through roots? J. Exp. Bot 49, 775-788.
- UNESA, (2005). Forestación y Reforestación. Sumideros de Carbono. En: Metodologías para la Implementación de los Mecanismos flexibles de Kioto Mecanismo de Desarrollo Limpio en Latinoamérica. Rivero Torre, Pedro, León, Gonzalo, Eichhamer, Wolfgang, Deputy, Gázquez Mateos, José Luis, González Santaló, José Miguel, Ferrando Bravo, Gerardo, Cisneros Gárate, Pablo, Pérez Pallarés, Diego. Capitulo 8.
- Watson, R. T., L. G. Meiro Filho, E. Sanhueza, A. Janetos, (1992) in Climate Change 1992The Supplementary Report to the Intergovernmental Panel on Climate Change Scientific Assessment (Cambridge Univ. Press, New York, 1992), pp. 2546.
- Winter C.K., Davis S.F. (2006) Organic foods, J. Food Sci. 71, R117R124.
- Wopereis M.C.S., Tamélokpo A., Ezui K., Gnakpénou D., Fofana B., Breman H. (2006) Mineral fertilizer management of maize on farmer fields differing in organic inputs in the West African savanna, Field Crop. Res. 96, 355362.






















